5 Fundamentals of Ecology
Jason Kelsey
As we saw in Chapter 1, the relationship between organisms and their surroundings is of great interest to environmental scientists. Here we will take a closer look at the ways the living things in an environment affect each other—that is, we will explore the science of ecology. Before we study dynamics involving groups, though, we will briefly consider the needs of individuals, because it is the drive to meet nutritional requirements that ultimately pushes a single organism into interactions with others.
Key concepts
After reading Chapter 5, you should understand the following:
- How different organisms acquire the materials and energy they need to survive
- How organisms can obtain and use energy
- The hierarchy of organization of natural systems, ranging from individuals to ecosystems
- The roles that different organisms play in ecosystems
- The distinction between ecosystem function and structure
- How biomes are defined and why they matter
- Various ways organisms interact with each other to fulfill their needs
- The rise and relevance of antibiotic-resistant bacteria
- Trophic levels and the movement of materials and energy through ecosystems
- The process of ecological succession and factors affecting it
5.1. WHAT IT TAKES TO SURVIVE
Despite obvious and dramatic differences in size, shape, structure, and lifestyle, every organism on Earth faces the same challenges: each must secure sufficient energy and materials if it is to succeed. We will soon see, however, that the biosphere features many different strategies to obtain necessary nutrients.
5.1.1. Energy
Demand for energy is high
Generally, organisms only carry out work that contributes to their basic survival. They struggle in the face of scarcity just to obtain the nutrients required to function each day, protect themselves, move (as appropriate), and stay alive long enough to produce offspring. The last item on that task list is extremely energy intensive and one of the most critical functions of living things. Put simply, current generations are driven to replace themselves with subsequent generations to ensure the continuation of their species (of course humans are not bound by all the same rules as other organisms, as noted in Box 5.1).
Box 5.1. Humans are not obliged to follow ecologists’ rules about energy and reproduction!
1. Energy. Humans must meet basic needs but are the only species that seems to find surplus energy for non-essential activities like video games, jogging, dancing, tennis matches, sex solely for the sake of pleasure, and Sunday drives.
2. Reproduction. Clearly, one can contribute to the continuation of a species without having offspring. But that discussion is beyond the scope of this book.
It is worth noting here that the reproductive strategies used to ensure the success of offspring and the continuation of a species vary. In some cases, for example, numerous fish and plant species, energy is devoted to producing as many individual progeny as possible rather than providing much (or any) parental care once the next generation emerges. On the other hand, organisms like humans concentrate their energy on raising one or a few offspring as a way to increase the likelihood that one’s genetic information survives into the future. Consult Box 5.2 for more details.
Box 5.2. How to best use your reproductive energy: overwhelming numbers or good parenting?
Most species allocate their reproductive energy in one of two ways. In the first, parents concentrate their energy on the release of large numbers of offspring—think clown fish, bacteria, frogs, and even dandelions (although the word is a bit clunky in some of our cases, we will assume “parent” is descriptive enough), and then do little or nothing to care for their young. This approach works because, although nearly all the new individuals are likely to die, enough survive to keep the species going. The second strategy involves the production of a few (or one) offspring that are protected and cared for to maximize their chances of survival. For example, relatively long-lived gorillas, elephants, orcas, cheetahs, and humans depend on intensive parenting (and all the energy it requires) to prepare their young before turning them out on their own. Ecologists employ a shorthand when referring to these two options: organisms using the first are known as r strategists, and those in the second group are called K strategists. What’s with the anti-intuitive letters? Suffice it to say that an equation employing several variables, including r and K, can be solved and yield either r- or K- dominant (more commonly called r- or K-selected) results. Finally, r and K are endpoints, and most organisms trend toward one of them. Note, though, that the reproductive strategy employed by some organisms does not fit neatly into the r / K classification scheme.
Phototrophs and chemotrophs obtain energy from different sources
Phototrophs. Light is the energy source for these organisms. Specialized structures in their cells allow them to transform solar radiation into chemical energy (note the suffix -troph is from the Greek for feeding—so, literally translated these are “light eaters”). Often, photosynthesis is the process by which these conversions are carried out (Chapter 4): using energy from the sun, atoms from inorganic molecules are linked together into the important products glucose and O2. The resulting molecules are stored in their cells until energy is needed to do work. Plants, algae, and some bacteria are in this group.
Chemotrophs. These organisms cannot harness solar energy but gain what they need through reactions involving pre-existing chemical compounds. Nearly all chemotrophs get their energy from organic molecules formed through photosynthesis or similar reactions, that is, directly or indirectly from phototrophs. Put into simpler terms, this group includes organisms that are not phototrophs: all animals, fungi, protozoa, and many bacteria.
5.1.2. Materials
Living systems are built of atoms
Recall from Chapter 4 that atoms can be bonded together in various ways to produce a dizzying number of different structures, including cells and tissues. Although many elements are required for an individual to be successful, organisms are classified into one of two groups according to their source of carbon (consult Box 5.3 for more about the elements that are most important to members of the biosphere).
Box 5.3. The ingredients for life
It would be fair to say that all living things are largely constructed of the same basic building blocks. As you might imagine, though, the amount of carbon, nitrogen, hydrogen, oxygen, phosphorus, and other elements present in cells varies among organisms. For example, bacteria, marine fish, land mammals, grasses, and trees need different relative amounts of carbon and nitrogen, but they all must obtain and concentrate large amounts of these vital elements if they are to be successful. Table 5.1 provides a very brief list of the nutritional requirements of two types of organisms, revealing both similarities and differences among members of the biosphere.
Table 5.1. Relative amounts of essential elements in bacterial and human cells. The numbers are the approximate percentage, by mass, of the total atoms present.
| Element | Bacteria[1] | Humans[2] |
|---|---|---|
| Oxygen | 20 | 65 |
| Carbon | 50 | 18 |
| Hydrogen | 8 | 9 |
| Nitrogen | 14 | 3 |
| Phosphorus | 3 | 0.5 |
| All others | ~5 | ~4.5 |
Autotrophs and heterotrophs use different sources of carbon
Autotrophs. When the prefix “auto” is combined with “troph” the resulting word can be taken to mean “self feeder.” Obviously these organisms are not able to conjure up carbon out of nowhere, so this literal translation overstates their powers. They do have an ability that most organisms do not possess: autotrophs produce useable carbon molecules from biologically unavailable carbon precursors. In other words, using carbon fixation, they can generate glucose from carbon dioxide they obtain from the air or water in which they live. Plants, algae, and some bacteria are autotrophs.
Heterotrophs. These “other feeders” are not able to fix their own carbon. They can only use organic, biologically available compounds. Heterotrophs therefore consume the glucose produced by autotrophs. Most species are in this group, including animals (like humans, fish, and insects), fungi, protozoa, and most bacteria.
5.1.3. Defining properties: energy and carbon sources
Many other characteristics affect survival and success, but we can say a lot about an organism if we know how it obtains energy and carbon. Using that information, we divide the biosphere into four basic groups.
Photoautotrophs
As with all the terms on this list, this one combines the prefixes related to energy and carbon source we saw in the past few paragraphs to create one descriptive word. Organisms in this category use the sun as their energy source (i.e., are phototrophs) and can fix CO2 into glucose (i.e., are autotrophs). Land-based plants, algae, and certain bacteria are photoautotrophs. They can succeed in many environments, both terrestrial and aquatic, as long as light is available at least some of the time (carbon dioxide, their source of carbon, is found in both air and water).
Chemoheterotrophs
These organisms use existing chemical compounds for energy and biologically available compounds such as glucose as their carbon source. Most of the biosphere falls into this group. Animals (e.g., insects, humans, rodents, fish), fungi, protozoa, and most bacteria are chemoheterotrophs. Since they do not use the sun directly, they can live in light or dark places provided useable carbon is accessible to them.
Photochemotrophs
A relatively small number of specialized bacteria use light energy but cannot fix their own carbon. Again, we can conclude that they must live in areas that receive some sunlight. Appropriate pre-formed carbon compounds—typically products and remains released by other organisms—must be present as well, though. These unusual organisms are usually found near the surfaces of bodies of water.
Chemoautotrophs
This last category, like the previous one, is made up of a comparatively small number of members. It consists of bacteria that need pre-existing chemical energy but fix inorganic carbon into glucose. Note that most chemoautotrophs do not use organic carbon compounds such as carbohydrates for energy, instead, inorganic compounds serve as their energy source. The sulfur-dioxide-eating bacteria mentioned near the beginning of this chapter are, arguably, the most famous (among those who keep track of such things) members of this group. In fact, the notion that chemoautotrophy could sustain life was only an unsupported hypothesis until the 1970s, when these organisms were discovered thriving in the complete absence of light near seafloor volcanoes.
5.2. ORGANISMS INTERACT TO MEET THEIR NEEDS
Now that we have been introduced to the basic needs of individuals, we are ready to study the many types of interactions seen within and among groups. Again, as we proceed through this discussion, you should not lose sight of the reason an organism operates as it does: it must fulfill the requirements presented in the beginning of this chapter if it is to survive. Put another way, the behaviors we will explore below only occur out of necessity. Non-human organisms do not act out of malice, greed, compassion, or other such impulses.
5.2.1. The hierarchy of ecology
One way to model ecological systems is with an inverted triangle. As is illustrated in Figure 5.1, complexity increases from bottom to top, with several different types of interactions possible at the top three levels.
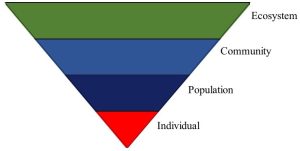
Individual
We know from Chapter 4 and elsewhere that an entire living being is hardly the least complex entity in the universe. Nonetheless, an individual is the simplest unit we will recognize as we begin to consider interactions. The list of possible examples would of course be extremely long, consisting of many millions of names of organisms. For our purposes, we will use one red-tailed hawk to represent this first level in our hierarchy.
Population
The next level in our scheme is the first one in which we start to see interactions. A population is defined as a group of individuals that are all members of the same species, that is, they can breed with each other to yield fertile offspring (see Box 5.4 for more about the species concept). In addition to reproducing, members of a population also can cooperate or compete. Note that this term can be applied on different scales: in principle, it includes every individual of a species on Earth, but it is not uncommon to identify a small subgroup in an area as a population of interest. For example, all the red-tailed hawks living and directly interacting near a mountain in, say, the state of New York, U.S.A., might make up a population studied by ecologists.
Box 5.4. Horses and donkeys and bacteria (oh my?)
The definition of species is one of the more straight-forward ones in ecology. Two individuals that can mate and produce fertile offspring are, by definition, members of the same species. It is therefore simple to objectively group organisms in this way. How similar they appear, how much they have in common genetically, and other potentially subjective criteria about whether they resemble each other sufficiently do not come into play. So, although a dog and a cat may share many characteristics (four legs, fur, tail, claws, and so forth) their inability to breed with each other puts them in separate groups. What about horses and donkeys? As you may know, they are closely related—so much so, that a union between a male donkey and female horse produces a mule. However, since these offspring are sterile (i.e., cannot reproduce), they are not members of the same species. It is all very logical and pleasing…until we consider bacteria. It turns out that the species concept only neatly applies to organisms that reproduce sexually, that is, where two parents combine their genetic material to produce one new individual. As we will see in Chapter 6, bacteria and some other microorganisms are asexual: a single cell divides into two progeny that are identical to the original cell. Further complicating things, bacteria are not particularly different in their shapes and appearances (i.e., there are many more species than there are different physical forms; Chapter 3). So they sort of look alike, and the offspring test is not applicable. How then can we make sense of this diverse group? Scientists have been wrestling with the problem for quite some time. When bacteria were first identified, microbiologists had to fall back on a comparison of physical and chemical characteristics, an approach fraught with subjectivity and uncertainty. During the past few decades, methods that allow for an objective analysis of DNA have evolved, making genetic relatedness increasingly important as a way to categorize organisms. Research in this area is ongoing.
Community
A group of populations that interact directly within a given space make up a community. Returning to our example, all the hawks, other birds of prey, songbirds, rodents, grasses, trees, and other organisms found on that New York mountain would be a community.
Ecosystem
This term is used by many ecologists to describe a level that encompasses the others in our hierarchy. It consists of all biological, physical, and chemical components active within a defined space. So, we would add the soil, air, and water—the relevant physical components and their properties—to the list of organisms imagined in our community above to define an ecosystem.
Why ecosystems matter. This, the most complex entity on our list, is the minimum unit necessary to sustain an individual, the simplest entity on our list. As we have seen previously, organisms are inextricably linked to their environments; they neither exist nor succeed in isolation, and they interact with their surroundings to fulfill their requirements. If an ecosystem fails to provide adequately, the individual organisms depending on it will fail as well. Furthermore, humans derive benefits from what are termed ecosystem services, including habitat for valued organisms, primary production, food, and important contributions to the hydrologic cycle.
Characteristics of ecosystems. In principle, Earth is one large ecosystem, although it usually is conceived and studied as a collection of many distinct ecosystems. For each, two important characteristics can be identified.
Although it has a different meaning outside of ecology, we will use this term to refer to the identity of the organisms present in a given ecosystem. Structure varies among ecosystems. For example, it is likely unsurprising that the community in a desert in northern Africa is not the same as that in a tropical rain forest in South America, the ocean near northern Australia, or the tundra in Russia. Since environmental conditions are so different in these places, so too will be the survival strategies used by organisms living in them. However, even in locations with similar conditions, such as the deserts in North America, structure will likely differ as well. As we know, environmental conditions affect organisms—it does not take much variation in temperature, moisture, or other properties to alter the organisms dominating an area.
This term refers to the processes active within an ecosystem. Unlike structure, this characteristic does not vary much from place to place. Regardless of the specific organisms present, the following processes or functions are active in all ecosystems.
Primary production. As we saw near the beginning of this chapter (as well as in Chapter 4), certain organisms are able to convert unusable carbon compounds into forms that are accessible to all members of the biosphere. Put another way, they produce fixed carbon for themselves and others. Remember that these autotrophs are almost always phototrophs as well: so, they also convert biologically unavailable energy from the sun into biologically available chemical energy (see Box 5.5 for a note about dark environments).
Box 5.5. What about places where the sun doesn’t shine?
Note that in those systems lacking sunlight, primary production is carried out by chemotrophic bacteria (see the early part of this chapter and Box 4.5). They are indispensable because of their ability to take a non-biological source of energy—in their case, that in inorganic chemical compounds like hydrogen sulfide instead of solar energy—and turn it into a biological form.
In terrestrial systems such as forests, deserts, and prairies, photoautotrophic plants such as trees, cacti, and grasses carry out primary production, respectively. This critical function is typically the work of photosynthetic algae and phytoplankton in oceans, lakes, and other aquatic ecosystems (plankton are generally small, free-floating organisms). Figure 5.2 shows examples of primary producers.

Despite the dramatic diversity in form, they share some common traits. Notably, their green color is associated with the machinery necessary to carry out the same function: photosynthesis. In other words, although adapted to very different environments, they all serve as primary producers for the ecosystems of which they are members.
Consumption. Recall from the discussion of the carbon cycle (Chapter 4) that carbohydrates and other organic forms of C move within the biosphere as organisms eat—consume—each other. This function is necessary in all healthy ecosystems as it provides chemoheterotrophs with the nutrients they cannot produce themselves and ultimately leads to the recycling of elements after the consumers die (see the next function on this list). The number of consumers on Earth is enormous, so coming up with examples is not difficult. On land, herbivores (plant eaters) such as zebra and deer and carnivores (meat eaters) like lions and wolves are among the organisms that carry out this function (Figure 5.3.) (more about herbivores and carnivores is presented below). Many microorganisms, including protozoa, fungi, and certain bacteria, are also consumers.

In water-based ecosystems, consumers range from microscopic zooplankton (they eat phytoplankton) to small fish, seals, tuna, sharks, and dolphins (Figure 5.4).

Decomposition. This third function is carried out by specialized consumers that eat the remains and waste products of organisms. Through it, nutrients are released and recycled. Decomposers also ensure that dead organic matter (Chapter 4) does not accumulate to any significant extent. Fungi and certain bacteria (distinct from the producers and consumers noted above) are responsible for much of the decomposition that occurs both on land and in water (Figure 5.5).

Of course, as we know by now, the specific decomposers found in different ecosystems must different—that is, the structure of decomposers varies even as the function they perform is consistent. For example, places that lack oxygen feature anaerobic decomposers whereas aerobic decomposers are found in oxygenated environments. Related to decomposers are organisms known as detritivores (detritovores are distinct from decomposers because the former are large enough to ingest distinct clumps of decaying material whereas the latter are so small that they act on and break down materials at a microscopic level). They feed on small organic fragments—known as detritus—of dead tissues and excrement. Terrestrial earthworms and some insects as well as ocean crabs and shrimp are good examples of these types of organisms (Figure 5.6).

Habitats and niches. These two terms denote distinct ecosystem properties. Habitat refers to the location in which an organism lives, whereas niche refers to the function an individual carries out—the role it plays, how it survives—in its ecosystem. Consider the three simple examples below, and note the difference in the way the words are used.
Great white shark
This fish lives in the coastal waters off the state of California, U.S.A. (its habitat) where it consumes large mammals such as seals (part of its niche).
Oak tree
This large plant grows in a forest in New Jersey, U.S.A. (its habitat) where it provides nesting opportunities for birds and squirrels, acorns for squirrels to eat, and is a primary producer (its niche).
Mosquitoes
These insects live near a lake in the African country of Kenya (their habitat) where they consume the blood of mammals and serve as a food source for bats (their niche).
Biomes. Simply put, a biome can be thought of as a type of ecosystem, an area characterized by a particular set of living and non-living features. Ecologists have identified several different biomes on Earth, defining and distinguishing them on the basis of climatic factors such as prevailing temperatures and abundance of precipitation as well as the types of dominant organisms (generally, the primary producers) present. The details of this topic are unnecessary for us, but future discussions (below and in Chapter 6) will benefit from an understanding of some essential concepts related to biomes.
How many biomes are there?
Unfortunately, the answer depends on who you ask. Some ecologists prefer to define a small number using very general criteria whereas others have established a larger number of narrowly defined biomes. In that context, it would be pretty safe to say the answer is in the range of five to fifteen.
What are Earth’s biomes and what organisms live in them?
Clearly the response to the first biome question suggests a complicated and variable answer is likely to be found here as well. We will keep things appropriately simple for our purposes. The two broadest types are aquatic (water) and terrestrial (land). Beyond that distinction, Earth’s major biomes are presented in Table 5.2.
Table 5.2. A sampling of biomes and their defining features[3].
| BIOME TYPE#, NAME[4] | DEFINING CHARACTERISTICS | DEFINING ORGANISMS | SAMPLE LOCATIONS |
|---|---|---|---|
| Aq: Marine | High salt content, wide temperature range | Algae, fish | Atlantic and Indian Oceans |
| Aq: Freshwater | Low salt content, wide temperature range | Algae, fish | Hudson River (U.S.A.), Nile River (Sudan) |
| Ter: Tropical rainforest | Rainiest biome, fewer than four seasons, warm all year | Many trees (not fir), ferns, flowers, monkeys, birds, insects | Brazil, India, Madagascar |
| Ter: Temperate rainforest | Second rainiest biome, experiences all four seasons | Many trees (including firs), rodents, birds, insects | Pacific northwest of North America, Coastal Chile, Britain |
| Ter: Temperate forest | Third rainiest biome, experiences all four seasons | Many trees (mostly leaf, not needle), rodents, bears, birds, insects | Northeastern U.S.A., Southern Canada, China |
| Ter: Grasslands | Generally less rainy than temperate forest, hot, dry summer, cold wet winter | Grasses and animals that eat grasses | Midwestern U.S.A., Argentina, Russia |
| Ter: Savannah | Intermediate between forest and desert | Grasses, trees, lions and their prey | About half of Africa |
| Ter: Desert | Second driest biome, can be very hot in the summer, cool in the winter in equatorial regions (very cold winters in polar regions) | Cacti, sagebrush, small mammals, reptiles, birds (polar: algae, grasses, penguins) | Southwestern U.S.A., Ethiopia, Antarctica
|
| Ter: Tundra | Coldest and driest biome | Algae, shrubs, caribou, polar bears | Alaska (U.S.A.), Siberia (Russia) |
Some words of caution should be offered before we move on. First, note that the table is not an exhaustive list, but a sampling for the purposes of illustration. A number of other biomes and characteristics could be included if we had additional space. Second, you should realize that, although some of the same names are used to indicate organisms present in different areas, species present vary considerably among biomes. For example, the term “birds” is used several times. As knowledgeable ecologists we know that only birds adapted to the narrowly defined environmental conditions in an ecosystem will survive there. So, the birds in a tropical rainforest will surely differ from those in a temperature forest.
Can the same biome be found in more than one place on Earth?
If you think of a biome as a type of ecosystem you will conclude that the answer to this question must be “yes”. Additionally, a further examination of the final column in Table 5.2 indicates that these types of ecosystems can be found in many places around the world.
5.2.2. Organisms can help or harm each other
Whether participants receive benefits or are inhibited is a fundamental and defining feature of the many possible interactions among individuals and groups. Three commonly observed interactions are briefly described here, although you should realize that these groups represent what might be viewed as end points on a continuum. In other words, the extent to which organisms are helped or harmed is not always easy to describe or categorize, and the boundaries among them can get blurred.
Competition
Everybody suffers. In this case, two or more individuals or populations are rivals for space, light, water, food or another resource that is in limited supply. Because none of them is able to access all of what it could utilize in the absence of competitors, all participants in this interaction are harmed. Natural systems are often characterized by scarcity, so organisms must develop effective strategies to obtain energy and materials in any case (what we have called adaptations previously). When demand for a critical nutrient exceeds supply, pressures on individuals and populations increase. If you recall how important the process of reproduction is to organisms, the reason competition brings on adverse consequences should become clear. Instead of devoting all its available energy to ensuring its genetic information is successfully passed to subsequent generations, an individual must expend precious nutrients in its struggle with others seeking to utilize the same resources. Note that competition does not necessitate or even imply direct confrontation among the participants, just that multiple parties inhibit each other through their efforts to meet their needs.
One ultimately wins. Ecologists have repeatedly observed that rivals for the same resources do not persist indefinitely in the same ecosystem. The competitive exclusion principle is an important concept—rule, really—that has developed as a result. In brief, it states: individuals of different species that have the same nutrient requirements cannot coexist. Put another way, they cannot occupy the same niche in the same space and time. What happens when multiple species try to access the same resources in the same place? The weakest competitor, the one least able to obtain what it needs, will disappear from that ecosystem. When this occurs, the winner, if you will, is unburdened by competition and will likely enjoy greater success as it can devote more of its resources to reproduction. On the other hand, the so-called loser could experience one of a few fates: become extinct (complete disappearance from Earth; see Chapter 6) if it is so specialized that it cannot exist elsewhere, adapt to occupy a different niche and exploit different resources in that ecosystem, or migrate to a location characterized by less intense competition.
Competition is common. It would be fair to say that competition is one of the most important forces shaping the living members of Earth’s numerous ecosystems, and this type of interaction is widespread. Given our space limitations, we will only consider a few good examples here.
Red Squirrels and Grey Squirrels
Aside from their obvious color differences, these two distinct species are strikingly similar (Figure 5.7).

They can live in the same habitats and consume the same food sources, but they cannot coexist without diminishing the success of one other. Observations made of populations sharing habitat in England have revealed the nature of their relationship and taught us a great deal about the importance of this type of interaction. The story gets somewhat convoluted, but in short: red squirrels are native, or endemic, to England, whereas grey squirrels are native to North America. Although these two species are equally successful when they are the only squirrels present in an area, red squirrels are currently being outcompeted by grey squirrels where they share habitat. Why do they overlap at all if the two species come from different continents? Well, back in the late 1800s humans intentionally imported them to Scotland, possibly for purely aesthetic and whimsical reasons. The two organisms never fought directly, but they became instant competitors. Unfortunately for the native reds, due to their higher rates of reproduction, more efficient use of available local food sources (abundant Oak Trees), and other advantages, the grey population grew quickly and spread aggressively. Now, many people fear that the red squirrels will disappear from the UK completely unless the grey squirrels can be controlled—something that has so far eluded us. Keep in mind that the competitive exclusion principle does not favor natives, just better competitors!
Black Walnut and other trees
The next time you take a walk in the woods, look for walnut trees growing in fields surrounded only by grasses (Figure 5.8). How is this possible? Black Walnut trees inhibit other trees that could encroach on their space and interfere with their success by releasing toxic chemical compounds into the soil. The poison affects other trees, but not grasses in an example of selective toxicity (Box 5.6, below, plus Chapters 9 and 15).

Two similar species of protozoa
Our final example comes from the unseen yet vibrant microbial world we explored in Chapter 3. Even though extremely small by our standards, organisms such as protozoa and bacteria interact in ways like organisms we can see with the naked eye. Similar to the squirrels above, Paramecium caudatum and Paramecium auerlia (two species of protozoa found in water—Chapter 6 presents more about the classification and naming of these protozoa and all members of the biosphere) are equally successful when grown in isolation and fed the same bacterial food source. However, when they occupy the same space, like a glass flask in a laboratory, both are slowed to some extent. Paramecium auerlia grows more quickly though, leading to the eventual disappearance of Paramecium caudatum; this is then followed by an increase in the success of the remaining, better-adapted, organism.
Mutualism
Everybody wins. Unlike competition, this cooperative interaction provides an advantage to all involved. Through different possible mechanisms, mutualism increases the success of participating individuals and populations. Under certain circumstances, these associations can even enable organisms to survive in environments they would otherwise find too hostile.
Mutualism is common. We are surrounded by cooperative relationships and even participate directly—generally unconsciously—in some ourselves. In fact, mutualism is widespread, affecting organisms throughout the biosphere. The specific ways organisms are connected are so diverse, however, that it would be difficult to generalize about them beyond the fact that they are mutually beneficial. A small number of examples will again have to suffice to illustrate this important and complex phenomenon.
Microorganisms and plants
Some bacteria can live within specialized structures of certain plants (mostly legumes such as alfalfa, clover, and beans) and fix nitrogen gas (N2) to a usable form of nitrogen such as ammonium (Chapter 4). The plant can then use the ammonium to produce its own proteins. In return, the plants provide fixed carbon for the resident N-fixing bacteria. The importance of this mutualistic relationship is hard to overstate as farmers take advantage of it to grow crops without the need to add nitrogen fertilizer to soil. We will return to the topic of fertilizers and soil in Chapter 9.
Microorganisms and cows
You likely have noticed dairy cows grazing in fields at some point in your life, but you may not have considered the complex mutualistic relationships they have developed with some tiny organisms (Chapter 3). Although they can ingest enormous amounts of grass, cows are utterly[5] dependent on the microbial community living within them to obtain nutrients from their food. In short, one of the chambers along the digestive tract of a cow, known as the rumen, is a very hospitable environment for certain bacteria and protozoa. Chemical bonds within the swallowed grass are broken by the microorganisms in the rumen, and the released nutrients are used by the cooperating symbionts. This association is critical because cows would otherwise starve to death—with full stomachs.
Microorganisms and humans
People are similar to ruminants in their reliance on bacteria and protozoa to help digest some of the food they ingest. Like cows, humans obtain nutrients from plant materials through the actions of specialized microorganisms living inside the intestines. Symbiotic organisms perform other services for us as well, including providing certain important vitamins and discouraging infection and disease by pathogens. For our part, we contribute nutrients and an appropriate environment for these microscopic partners. Review Chapter 1 for more on this relationship.
Deep-sea fish and bioluminescent bacteria
Many organisms living in deep, dark waters have developed mutualistic relationships with bioluminescent bacteria (i.e., they can convert some of their energy into light). In the case of the anglerfish, the bacteria live inside a long, thin structure, an esca, that is attached to the animal’s head and dangles near its mouth (Figure 5.9).

They receive nutrients and a suitable habitat, and in return light up and cause the end of that protrusion on the fish to glow. This arrangement provides a useful advantage for anglerfish: the esca serves as a lure for unsuspecting prey attracted to one of the only light sources in the dark water (hence the name “anglerfish”).
Fungi and algae grow as lichens
Our final example is one of the most famous and well-studied cooperative associations in ecology. In this case, the relationship is so highly developed and specific that two separate organisms, fungi and algae, appear to be a third entity, a lichen (Figure 5.10).

Because of the contributions made by each of the partners, lichens are successful in locations too hostile for fungi or algae to grow on their own. Algae are primary producers, providing fixed carbon and energy, and fungi have structures that enable them to obtain and hold onto water as well as other nutrients that may be in short supply. Lichens can grow on barren rocks, dead trees, and even the sides of buildings, and play an important role in the process of ecological succession (described in an upcoming section of this chapter).
Winners and losers, part 1. This is one of two types of interactions through which organisms derive benefits at the expense of others. Here, a parasite causes harm to its host by living on or inside that larger organism (the other case, predation, is described below). Although we will only consider a small number of examples of parasitism, keep in mind these types of relationships are varied and numerous.
Pathogenic bacteria and humans
We have seen previously that members of the group known as bacteria carry out an enormous number of vital functions and are indispensable to the biosphere. Those that are parasitic, though, can bring on diseases that run the gamut from mild to deadly. For example, strep throat, tuberculosis, syphilis, cholera, and pink eye are among the many conditions caused by bacterial infections. A few cells of the pathogen can enter a person, evade defense mechanisms, and rapidly increase in number inside the body system that provides the best environment for them (throat, eye, etc.). The details vary widely among different diseases, but in general terms the bacteria are parasites: they divert nutrients, inhibit normal functions, cause tissue damage, or otherwise harm their host. Now, humans employ some natural mechanisms to fight infection, ranging from non-specific responses like fever to cells that target specific invaders, but they are not always successful. What if body defenses fail to eliminate the parasite? Permanent damage or death may occur without medical intervention such as the administration of chemical antibiotics (see Box 5.6 for more). Viruses and other microbes can behave like parasites as well, with outcomes like those brought about by bacteria. Treatment of diseases they cause, however, is not the same.
Box 5.6. Antibiotics: winning a battle, losing the war, against pathogenic bacteria
We have been fighting disease-causing bacteria for as long as humans have been on Earth, but up until relatively recently, there was little we could do against many of them. People infected with pathogenic bacteria had to, for the most part, depend on their immune responses that are only partially effective against such organisms or on medical interventions that might treat or alleviate the severity of symptoms yet do little or nothing to actually eliminate the invaders and the diseases they cause. Then, in the 1920s, an important discovery changed the balance of power: penicillin, a chemical substance produced naturally by a certain type of mold, was found to destroy many harmful bacteria. Starting in about the 1940s, antibiotics gained wide availability and were able to cure diseases such as tuberculosis, cholera, syphilis, strep throat, and others that had killed so many people throughout history. The success of penicillin and the other antibiotics used since is attributed to a phenomenon known as selective toxicity. Put simply, different organisms can react very differently when exposed to the same substances. Although they have much in common as living entities, humans and bacteria are distinct in several important ways. For example, most pathogenic bacteria build cell walls outside of their cell membranes (Chapter 3) whereas animals lack such structures. Consequently, humans can ingest penicillin, which targets wall synthesis, to kill off bacteria that are making us sick without causing undue harm to ourselves (aside from people who are allergic to the drug). As brilliant as these medicines may seem, some serious adverse effects come along with their use. For one, in addition to killing pathogens, antibiotics can also kill mutualistic bacteria inside places like our intestines, making it temporarily difficult to properly digest food. However, a second phenomenon is far more problematic: the development of antibiotic-resistant bacteria. During the decades since we discovered penicillin, many populations of bacteria that were once vulnerable to antibiotics are no longer killed by such chemicals. In practical terms, this means you might effectively treat strep throat with penicillin a few times but then discover the drug has no effect on a subsequent bout with the disease. Because bacteria can quickly evolve a way to overcome the adverse effects of the drug, they continue to live and cause harm to their host. How? Recall that organisms are affected by the stressors they encounter in their habitats. To be blunt, a species will either fail (die or at least be forced out) or adapt a survival strategy and persist. An antibiotic introduced into any system will kill only susceptible bacteria, leaving the few unaffected cells to reproduce with reduced competition. Once their numbers increase, these resistant individuals can continue to cause disease unhindered. We have a lot of evidence of this phenomenon in public-health records, but it can also be witnessed in real time. Laboratory experiments have repeatedly shown how a population of bacteria exposed to an antibiotic can change from sensitive to resistant in a matter of days (more in Chapter 6).
Should we be concerned about antibiotic-resistant bacteria? The short answer is: yes. As a population of pathogens becomes resistant to an individual antibiotic, a new chemical that targets different sensitive structures in the bacteria may be employed to fight a disease (e.g., instead of penicillin to block cell wall construction, you use tetracycline because it disrupts other prokaryotic processes). Unfortunately, the response to that second antibiotic will be as it was for the first, yielding a population that is now unaffected by two different drugs. A third and a fourth, and many other antibiotics can be tried, but the outcome is always the same. Certain pathogenic bacteria have even become resistant to every known antibiotic. One instance of multi-drug resistance recently identified by public health officials is a strain of bacteria that causes the common sexually transmitted disease gonorrhea. This once readily treatable condition now could potentially persist in infected individuals, causing many long-term health problems. A return to the situation that prevailed prior to the discovery of penicillin, that is, we possess no effective defense against bacteria, may well occur in the not-so-distant future.
Can we do anything about antibiotic-resistant bacteria? The short answer is: not much. Arguably we have vastly increased the pace of an inevitable process by the way we have used antibiotics, but it may be impossible to prevent in any case. We could probably slow the spread of antibiotic resistance among all of Earth’s bacterial populations by taking the following steps.
1. Use far fewer antibiotics. Keep in mind antibiotics are not effective against all microorganisms. For instance, viruses and protozoa are insensitive to penicillin and other drugs that target bacteria. Public health officials note that many physicians, particularly in the U.S., overprescribe antibiotics, using them to combat conditions that will generally run their course without chemical therapy. So what’s the harm in being extra cautious? Well, a large amount of those antibiotics we ingest exit our bodies in urine and enter natural water systems. Thinking like an environmental scientist, you might picture what happens when populations in soil and water are exposed to antibiotics: they adapt. Put somewhat differently, as we use more and more of these chemical substances, the very organisms we are trying to kill gain more and more of an advantage.
2. Develop new antibiotics. This step may seem rather straight forward and obvious. In principle, we might be able to keep better pace with bacterial evolution by quickly and regularly designing new drugs, but there is currently little effort being devoted to this goal. The reasons for this apparent lack of interest are numerous. Suffice to say, the outcome of it could include an enormous public health crisis in the coming years.
Some have called it an arms race, and humans have gained only a temporary advantage in it.
Tape worm inside a dog
Anyone who has ever lived with dogs knows that these animals will eat nearly anything that they perceive might be food—roadkill, excrement (their own or others’), debris they remove from their own bodies, many inanimate objects—with gusto and seemingly no concern about the potential consequences. Unfortunately, among other problems arising from their indiscriminating palate is the tendency to ingest fleas and the tapeworm larvae they can carry. These parasites grow inside the digestive system and cause harm by eating some of the nutrients consumed by the dog. This association between the two organisms can last for a relatively long time, as it is generally not all that dangerous to the animal (medical intervention will kill the worm). Incidentally, humans are also susceptible to tapeworm infestation. In some cases, people have been known to intentionally ingest the parasite in the interest of weight loss[6].
Wasp larvae and tomato hornworms
One of the scourges on tomato growers is a green caterpillar known as the tomato horn worm. These insects can eat the leaves of the plants and destroy crops. Interestingly, this pest is itself subject to a parasite. In short, a certain type of wasp will lay its eggs on the skin of the hornworm. The larvae that hatch from the eggs slowly eat the caterpillar until they emerge as adults. While they are feeding and developing, the larvae can be seen protruding from the back of the still-living hornworm (Figure 5.11). This relationship ultimately ends in the death of the host, unlike some of the parasitic relationships described above.

Predation
Winners and losers, part 2. Predators are like parasites in that they obtain what they need while bringing about some level of harm to another organism. However, this relationship differs from the former in three critical ways. First, parasites tend to be small relative to their hosts whereas predators are larger than, or at least capable of overpowering, their prey. Second, the length of time parasites can interact with their hosts is almost always longer, ranging from days to years, than that of predators and their prey, which is in the range of minutes. Nutrients stored in prey organisms move to predators, as the former are killed and consumed by the latter (more about this flow of nutrients within ecosystems is described in the next section). Third, although predation is almost always fatal to the prey, parasitism need not kill the host (think dogs and fleas). One final point bears consideration: generally speaking, the term “predator” is reserved for organisms that eat other heterotrophs like animals or certain microorganisms (animals that eat the flesh of other animals are also known as carnivores). Plant-eating consumers tend to be called herbivores to reflect the fact that they eat plants or other plant-like cells. A small number of organisms, humans, as well as certain rodents and insects for instance, are appropriately classified as omnivorous because they can eat both animals and plants.
Predators eat to survive. Consumers get energy and materials by eating other organisms. Recall from earlier in this chapter that, although they could appear to be violent or cruel when viewed through the lens of human emotions, predators simply act in response to their fundamental needs. Predation is active throughout the biosphere, but as before, we will only briefly consider a few examples of it.
Spiders consume insects
A number of spider species build webs to catch unsuspecting insects. Once ensnared, the prey is wrapped up by the predator to be eaten later (Figure 5.12).

Orcas consume seals
Killer whales, also known as orcas, are large predatory animals which eat other animals such as salmon, seals, and tuna (Figure 5.13). Those prey species are themselves predators on smaller organisms. We will examine feeding relationships in ecosystems, known as trophic interactions, in more detail shortly. The role that predation has on the movement of pollutants in ecosystems is another important topic, one we will cover in Chapter 15.

Protozoa consume bacteria
Predation is a way of life at all scales, from the familiar visible animals noted here to the microscopic world we explored in Chapter 3. Like orcas, spiders, lions, wolves, and so forth, protozoa are consumers that obtain nutrients through ingestion of other, smaller, organisms such as bacteria. In fact, a single protozoan can eat thousands of individual bacterial cells during its life cycle. This predation, or grazing, plays an important role in many ecosystems because it can control the numbers of bacteria present and can serve as the basis for many ecosystems.
5.3. MATERIALS AND ENERGY MOVE THROUGH ECOSYSTEMS
As we learned in earlier parts of this chapter, individuals must fulfill specific needs to survive. Although a variety of strategies can be used, most often materials and energy are obtained from other organisms. Put simply, producers obtain nutrients and convert them into biologically available forms, and then consumers live off of those producers directly or indirectly.
5.3.1. Hierarchy of feeding relationships
Trophic levels
Organisms are grouped according to the number of steps they are from the original source of energy (typically the sun, as we saw above) for a particular ecosystem. Each of these groups is called a trophic level (Figure 5.14).
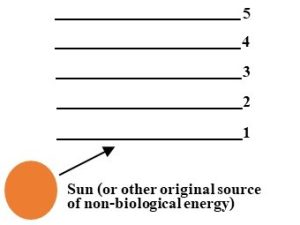
For example, since they obtain their energy directly from the sun, photoautotrophs such as plants and algae, as well as all other primary producers, are in the same category: the first trophic level. Organisms that consume those at the first are in the second trophic level and are themselves consumed by predators at higher levels. A simple, idealized representation of the arrangement and characteristics of trophic levels is presented in Table 5.3.
Table 5.3. Characteristics of different trophic levels. In some cases “RANKING / NICHE” provides more than one possible term for organisms in a given level to suggest different ways to view their roles.
| TROPHIC LEVEL | RANKING / NICHE | DIRECT ENERGY SOURCE | EXAMPLES |
|---|---|---|---|
| Fifth | Quaternary consumers, Tertiary carnivores | Organisms at the fourth and likely third and second trophic levels | Eagles, sharks |
| Fourth | Tertiary consumers, secondary carnivores | Organisms at the third and likely second trophic levels | Foxes, seals |
| Third | Secondary consumers, primary carnivores | Organisms at the second trophic level | Songbirds, small fish |
| Second | Primary consumers, herbivores | Primary consumers | Caterpillars, zooplankton |
| First | Primary producers | Non-biological energy source like the sun | Plants, phytoplankton |
Note that organisms are ranked on the basis of the highest level they could possibly occupy. So, in the table, the eagle is placed in the fifth trophic level because it can feed on organisms in the fourth trophic level (e.g., the fox). However, it may skip down to a songbird in the third trophic level if presented the opportunity to do so. Since few organisms can eat both plants and animals (i.e., are omnivores, as define above), the diets of high-level consumers tend to be restricted to prey in the second level and above.
Food webs
One useful way to visualize trophic relationships is through a diagram known as a food web. This kind of graphical representation shows the possible interactions among trophic levels, or put somewhat whimsically, can be used to answer the question “who eats whom?”. Importantly, our model is not a chain, but a web. Although people commonly use the term “food chain” in reference to trophic interactions (and even outside the realm of ecology as a metaphor in any hierarchical system, e.g., “since she is higher on the food chain, she makes more money”), a linear structure vastly oversimplifies the situation. Figure 5.15 enables a comparison between the use of chains and webs in this context.
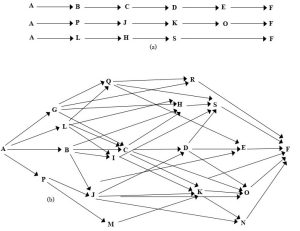
Note how chains necessarily and unrealistically suggest that each organism has only one source of food and one predator, whereas a web more accurately presents the complex and varied nature of the movement of materials and energy from lower to higher trophic levels. Put another way, one organism can eat and be eaten by a number of different other organisms. The food web modelled by Figure 5.15 implies six possible trophic levels (A is at the first, F is at the sixth), although a careful examination of it reveals that energy does not always move through six levels from organisms A to F. In fact, multiple possible pathways exist (examine the figure to see for yourself, reading left to right). In any case, the organism that occupies the highest trophic level, the one that is not ordinarily a source of food for any other, is referred to as an apex predator (more below). A diagram of an example food web studied in the Chesapeake Bay is shown in Figure 5.16.
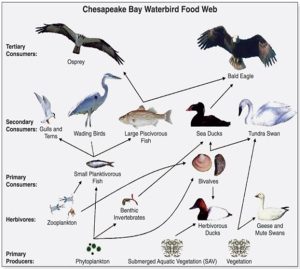
The number of trophic levels and the complexity of the webs can vary considerably among ecosystems. Broadly speaking, extreme, barren, or ever-changing environments are characterized by fewer organisms and simpler models. In a very dry desert, for example, it is more difficult to develop proper adaptations and survive, so there are fewer different species present. Put another way, there is low biodiversity in such an environment Trophic interactions are therefore relatively simple in such places. On the other hand, a more hospitable, stable, or nutrient-rich habitat such as a coastal ocean or tropical rainforest tends to support a much wider variety of species and has a more complex food web as a result.
5.3.2. How many trophic levels are possible?
On first pass, this question might appear to be inconsequential, one that is a simple matter of book keeping suited for academic discussions only. However, the answer is rooted in some fundamental principles we studied in Chapter 4 and is critical to a discussion of ecosystems.
Available energy decreases with increasing trophic level
We learned previously in this chapter that energy is converted into a biologically available form by members of the first trophic level. In serving as food sources, these primary producers then supply energy to higher-level consumers. The amount of usable energy that can pass from one level to the next is quite low, though, because most of it is used to do work or passes to decomposers before consumers can access it. You might be surprised to learn that somewhere between 90 and 99% of the total energy entering a trophic level is lost—that is, nearly all of it becomes unavailable by the time organisms at the next level can access it. This phenomenon can be visualized with a graphical representation known as an energy pyramid (Figure 5.17).
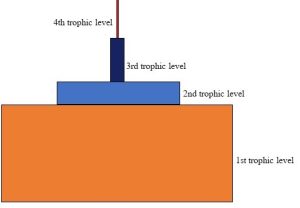
Just where does this lost energy go? Two important processes divert energy away from consumers, bringing about crucial consequences to all ecosystems.
1. High-grade energy is degraded to heat. The laws of thermodynamics we first encountered in Chapter 4 are ever present, governing ecological and all systems. Organisms at each trophic level use the relatively high-grade chemical energy to which they have access for work such as reproduction and movement. In the process, that energy is degraded to heat, a form that is unusable by higher-level consumers. Picture, for example, the feeding relationship between a gazelle and a cheetah. The prey species is an herbivore, obtaining necessary nutrients by eating grasses. The predatory big cat gets what it needs by eating the gazelle, but it must expend a substantial amount of energy hunting and killing its prey. For its part, the gazelle utilizes stored energy to power all its functions, including running for its life when pursued by a cheetah. A protracted chase will result in that much less available energy for the successful cat, and its net gain will be a function of how much both it and the gazelle expended. The prey may or may not elude its predator, but there is no escaping the second law!
2. High-grade energy passes to decomposers. In addition to the work and associated conversions described in the previous section, various other types of losses also remove usable energy from a particular organism and trophic level. As we saw in our discussions of ecosystem functions in the early parts of this chapter and the carbon cycle in Chapter 4, waste products released by organisms contain a lot of high-grade chemical energy. It is not just excrement, though: plants also lose branches, leaves, bark, and the like, and animals shed hair, skin, and other body parts during their normal cycles. Decomposers are adapted to extracting energy stored in these waste products and using it to enable the work they must do. Ultimately, low-grade heat energy is released as those fungi and bacteria grow and reproduce.
Declining available energy limits growth at high trophic levels
The dramatic drop in available energy just described greatly restricts the number of trophic levels present in any ecosystem. In fact, no more than approximately six consumer levels are possible in most food webs, with fewer seen in many situations. The amount of energy required to support yet another consumer level above the current apex predator simply exceeds that which is available to do so. Furthermore, the total amount of living material, termed biomass, in each level declines quickly and dramatically with distance from the original energy source (Figure 5.18).
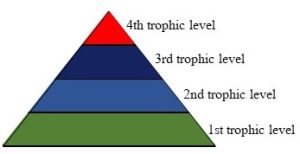
As counter intuitive as it might seem, in terms of mass, there are far more primary producers than there are herbivores, more herbivores than primary carnivores, and so forth up to the apex, because of the massive energy inefficiencies described above. Yes, higher-level animals do have a higher concentration of stored energy in their tissues than do plants (that is, amount of energy per gram of flesh), but the total amounts of both mass and energy in a given trophic level are far lower than those in the levels beneath it.
5.4. ECOSYSTEMS DEVELOP AND CHANGE IN RESPONSE TO STRESS
The final section of this chapter will address two fundamental questions about ecology we have yet to ask: first, how do ecological communities come into being and, second, what happens to an ecosystem that is damaged or otherwise subjected to stress?
5.4.1. Ecological succession changes ecosystem structure with time
Contrary to what might be our instincts or expectations, a system such as a mature forest did not simply emerge fully formed at the beginning of time and remain stagnant since. Instead, it and all ecosystems developed through ecological succession, a dynamic process by which the organisms dominating an area are replaced sequentially. Using a term we encountered earlier in this chapter, we could also say succession changes the structure of an ecosystem with time. These changes continue until a final (really long-lived) stage is reached (see Figure 5.19 for an overview).
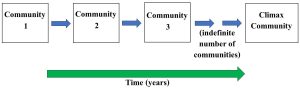
Importantly, unlike the earlier communities, the climax is not easily replaced but persists until conditions change sufficiently to disrupt it (more below). We will see shortly that the concept of ecological succession is not nearly as straight forward as the idealized model described here. We devote space to it, though, because it is a fundamental idea in ecology that can be used to study and predict how ecological systems develop, change, and respond to stress.
Ecologists recognize two types of succession
The two types are distinguished on the basis of how much destruction precedes them. Primary succession follows a severe disturbance, one that obliterates a community. In other words, a new ecosystem is formed on a stark, barren landscape. Alternatively, secondary succession restores an existing community subjected to relatively minor damage. Here, recognizable remnants of the previous ecosystem are left behind, and succession can be viewed as a mechanism of recovery.
Primary succession. Many forces can produce barren environments, either by generating completely new habitat or by wiping out an existing community. A volcanic eruption, for example, is a powerful and transformative event; among other things, it can eject large amounts of lava onto Earth’s surface (more in Chapter 7). This extremely hot molten rock tends to engulf anything in its path—forests, grasslands, houses and so forth—burning and burying it. Once the lava has cooled, it becomes solid rock, devoid of any life. Other natural causes of this level of destruction include melting of large glaciers that have covered an area for thousands of years, and collisions with asteroids. Humans can also clear land through agriculture (Chapter 9), construction, pollution (Chapter 15), and even warfare. We will devote some more attention to the ways human activity affects succession near the end of this chapter. Whatever the force, primary succession can follow it, changing an apparent wasteland into an area characterized by a complex, productive, and thriving community of organisms. You should keep in mind that this process is active in any location subject to severe changes, be they terrestrial or aquatic, hot or cold, wet or dry, etc. The upcoming section (along with Chapter 6) presents more information about the different types of ecosystems found in different places.
Secondary succession. Unlike primary, secondary succession begins on a landscape that still includes organisms and visible sources of nutrients and other vital means of support. Events such as forest fires, floods, earthquakes, landslides, and hurricanes can damage but not obliterate existing communities and are among the precursors to secondary succession. Here too, anthropogenic sources of stress, activities like tree cutting, various types of harvesting, and others, can set the stage for this type of ecosystem response.
More on the relationship between the succession types. Note that the two types of succession we just defined are distinct processes and should not be understood to be sequential. That is, despite what you might assume from their names, primary does not precede secondary. Instead, they each represent different paths by which an ecosystem can progress toward a climax community. Yes, the mechanism of replacement is integral to both, and it can be reasonable to even think of them as starting at different points on the same continuum. But given the many possible variables affecting the trajectory of succession, including the nature of the destructive event that came before it, it is appropriate to keep them separate from each other.
Some clarifications and context
Several important assumptions require our attention before we proceed. First, when studying succession, ecologists generally focus on the primary producers present in an area. In other words, first-trophic-level organisms, and how they change with time, are used to define different stages of succession. We expect that the identity of consumers at the second level and above will be a function of the producers present and will also change accordingly. So, in a land-based habitat, we would only keep track of plants and plant-like organisms. Second, primary producers attempt to procreate by releasing seeds (or analogous structures). Through various mechanisms, for instance on the wind, seeds are carried from their points of origin (he distances travelled can be enormous, hundreds or even thousands of kilometers). Usually, the seeds land in places inhospitable for them to grow. On occasion, though, they fall into exactly the right place at the right time, finding an environment that is well suited for them. Keep this second point in mind as we explore succession because it is important to remember that communities will most likely not be restricted by availability of organisms, rather by lack of necessary resources to support growth. Third, an area that exhibits little or no obvious growth and contains no visible sources of nutrients (e.g., no soil) is termed barren for the purposes of our discussion. Finally, succession should really be thought of a continuum, but for ease of study, we separate it into discrete stages as described here.
Succession involves many different species and steps
As noted, we will view ecological succession as a series of replacements. Earlier-stage species, are replaced by later ones that are themselves subsequently replaced. Unsurprisingly, a community that takes over an area does so because it is well suited to live there. What we might not expect, though, is that the actions of those successful organisms can change the environment so much that it becomes a better place for other organisms—members of a later-stage community—to live than it once was. In other words, earlier-stage organisms set the stage for their own departure because they often improve conditions for later-stage organisms that otherwise would not have been able to survive there. Changes continue until a stable end point is reached, what we termed the climax community. An important feature of this phenomenon is that the organisms dominating the different phases of succession possess different adaptations.
Pioneer species. These organisms begin the process of succession, so, in the idealized diagram in Figure 5.19 we saw above, they would make up Community 1. They colonize areas such as barren rocks, sand, ocean floor, or others that lack an obvious existing community. Although most species will fail to grow under such harsh conditions, pioneers possess traits that enable them to reproduce and take over seemingly unwelcoming landscapes. Notably, they devote an unusually large fraction of their energy to reproduction, dispersing large numbers of seeds (or the equivalent) far and wide. In a suitable environment, the seeds will quickly grow into mature forms, release their own reproductive structures to start the next generation, and die after a relatively short life. Their fast and intense existence requires an enormous amount of available energy, generally in the form of abundant, direct sunlight. The adjective “opportunistic” is often applied to them for they are well adapted to take advantage of habitats few others can tolerate.
Climax community. Although this is not the second but the final stage, we consider it immediately after pioneers because it represents the other endpoint in the process of succession and provides us with an opportunity to compare the traits common in these species to those found in pioneers. It is not quite this simple, but it is broadly fair to say that organisms in the climax community are everything pioneer species are not. Relative to colonizers, these species devote less energy to reproduction, grow slowly, live longer, and require less direct exposure to sunlight. Table 5.4 summaries the major differences between these two types of organisms.
Table 5.4. A comparison of important traits of pioneer and climax species.
| TRAIT | PIONEERS | CLIMAX |
|---|---|---|
| Amount of energy devoted to reproduction | High | Low |
| Growth rate | High | Low |
| Length of life | Short | Long |
| Shade tolerance | Low | High |
| Likely to be replaced? | Yes | No |
Early-, mid-, and late-stage communities. Now we consider the species that dominate between the two end points described above. Even as pioneers colonize an environment, many kinds of seeds continue to blow into the area. This time, though, since the conditions have been changed somewhat, a different group of organisms begins to appear. It has much in common with pioneers—need for direct sun, rapid growth, short lives—but it is not adapted to succeed on a completely barren landscape. In addition, these early-stage successional species will likely be larger than pioneers. As time progresses, the pioneers are outcompeted, and the character of the community changes. What happens to those newcomers? Their presence will likely facilitate the growth of additional new species, termed mid-stage successional species, that eventually take over the area. Regarding the important traits we have identified, they are intermediate between pioneers and climax organisms. More time brings further alterations away from the earliest conditions, allowing for the growth and dominance of increasingly long-lived, shade tolerant, and large primary producers. Late-stage successional species have more in common with those in the climax community than they do with pioneers but are themselves slowly pushed out by the species that are best adapted to this final, hypothetical environment. You should bear in mind that, beyond the changes noted in the survival strategies used by organisms, the community itself can take on different characteristics as well. Notably, although this trend is not universally seen, some researchers have observed how the total number of different species present is generally low in the earliest communities, increases through the middle stages, and then may decrease again at the climax (more in Chapter 6). If we think about the traits that are most advantageous at the various stages, we might predict this trend. The ability to colonize a hostile area requires specializations that are not common in the biosphere. We learned earlier, for example, that lichens can grow in very nutrient-poor settings. Consequently, they are among the few species able to act as pioneers. As challenges presented by an environment become less severe, there is a potential for a greater biodiversity because the necessary traits are less unusual. A decline in the total number of species in late stages could be explained by the fact that such a community is crowded, nutrients are scarce, and sunlight is not as available as it once was. Again, the conditions require a somewhat narrow and specialized set of adaptations. Despite the apparent logic in this reasoning, though, many ecosystems defy expectations. Clearly the many variables affecting community composition warrant additional study.
Succession affects all environments
Ecological succession surrounds us, controlling the way natural systems develop. The small sampling of examples included here demonstrates that, although the process is fundamentally the same wherever it is active, the details vary considerably as a function of prevailing environmental conditions. Furthermore, succession within a biome (described in an early part of this chapter) often follows a predictable pattern.
Temperate forest.
Succession can transform a landscape left barren by a volcanic eruption
Note that the following steps represent an idealized view of what is possible, not what is inevitable in this biome. That is, some variations from location to location are likely.
Bare rocks support little or no life (Figure 5.20). Whatever is living on this cooling field of volcanic rock is not immediately apparent.

Lichens and mosses colonize the rocks (Figure 5.21). Seeds of many grasses, shrubs, and trees will land here, but due to the lack of soil and readily available nutrients, they will not grow. After a short time, the presence of pioneers starts to produce small amounts of soil. How does soil develop? Recall from our discussions of the carbon cycle and decomposition (Chapter 4 and early in this chapter) that some of the decomposed waste products and remains of organisms can accumulate in the lithosphere. In this case, they could fill small cracks in the rocks, fundamentally changing the character of this environment. We will see much more about soil formation in Chapter 9.

Grasses start to grow in the soil created from the remains of pioneers (Figure 5.22). The presence of even a small amount of soil, combined with the direct sunlight that would be available in the absence of any large organisms, allows these small organisms to quickly spread across the area.

Small shrubs, plants like milkweed, start to grow (Figure 5.23). The presence of the fast-growing grasses leads to the production of even more soil and a more welcoming habitat for slightly larger organisms.

Small trees like black walnut and birch can now grow (Figure 5.24). These early-stage organisms quickly mature and partially block some of the incoming sunlight. Soil and nutrients continue to accumulate.

Saplings of slow-growing trees such as maple and oak start to emerge (Figure 5.25). About this time, the early-stage trees from the previous step approach their maximum size and age and drop seeds to start the next generation. However, since the presence of shrubs and trees greatly reduces the availability of direct sunlight, seeds of the early-stage trees will struggle to grow.

The maple and oak trees get larger, reducing sunlight even further (Figure 5.26). The earliest-stage organisms are likely gone by now, as are many of the early and mid-stage trees. Seeds of the late-stage trees can grow in partial shade, and those species begin to take over the area. Often, such organisms are very straight and tall, growing densely enough that their crowns (i.e., the top sections containing the most leaves and branches) are in contact with each other. The resulting phenomenon is called a canopy, a largely continuous structure at the top of the forest. Like canopies built by humans, it blocks much of the incoming sun, creating a shady forest floor.

The climax community made up of tall maple, oak, and similar trees persists (Figure 5.27). Under idealized circumstances, these species will dominate and not be subject to replacement. In reality, a climax forest may not last particularly long—a number of factors and forces can disrupt it (as stated above; we will see more below as well).

How long does it take?
The time required for a forest to arise from a barren rock field varies, as you might expect. However, to provide a sense of scale, it would not be surprising if the entire process was completed within a century. In fact, the development of a field filled with grasses and small shrubs (review Figure 5.23) could occur after only a few years, depending on the specific conditions affecting the relevant environment.
Before we leave forest succession, we should recall one of the clarifications made earlier: succession is generally described according to the primary producers present in an area. We would expect the identity of animals to change from small insects, rodents, and birds in the early stages to larger mammals, larger birds, and small reptiles in the later stages. The climax forest could be habitat for deer, bears, lizards, owls, and so forth, animals that would be largely absent from open, grassy fields.
Barren sand can become a forest
Although not formally identified as a biome like the temperate forest we just studied, a coastal sand dune is a distinctive area characterized by unique conditions and challenges for members of the biosphere attempting to live there. Through the following process it can undergo succession from a seemingly lifeless mound to a teeming, if small, forest.
A barren sand dune is an unwelcoming habitat for most (Figure 5.28). Organisms struggle to live in such an environment for two important reasons. First, sand is quite nutrient poor so supplies little of what organisms need to survive. Second, sand is not stable, but moves due to wind and precipitation. Most plants cannot survive in such a harsh habitat.

Specially adapted plants withstand moving sands (Figure 5.29). Because they have extremely long roots—several meters is possible—certain grasses can colonize a dune. The roots anchor the plants and allow them to grow as the sand shifts. Even more importantly, though, the presence of the root network stabilizes the dune somewhat. Organic material also begins to accumulate as the remains of the grasses are decomposed.

Other grasses and small shrubs begin to appear. Thanks to the actions of the pioneers, the dune is a more hospitable habitat than it was: it is now both comparatively stable and nutrient rich. The addition of organic material also increases the amount of water available for plants (we will see much more about the factors affecting soil fertility and water retention when we discuss agriculture in Chapter 9).
The climax community is dominated by small trees (Figure 5.30). Since sand dunes can be located on beaches and coastlines all over the world, the specific identity of climax trees found on them varies enormously. In any case, fast-growing trees such as beeches, pines, and birches are likely to make up the stable community in these environments.

Not everything can grow on a dune. As we know from several previous discussions (e.g., Chapters 1 and earlier in this chapter) organisms that are best adapted will dominate an area. So, although both are terrestrial systems, the temperate forest supports a different set of organisms than does the sand dune. The climax community near the coast must withstand salt, wind, intense storms, and other stresses that an inland community would not face. We will return to this point shortly.
Freshwater pond.
An aquatic system becomes terrestrial
Succession within this biome can lead to dramatic changes, shifting from a community dominated by algae, fish, and other water-based organisms to one dominated by trees, rodents, and other land dwellers.
Nutrient accumulation can cause eutrophication
We have seen this phenomenon before, albeit in a different context, in Chapter 4. If you recall how excess nitrogen and phosphorus entering a body of water can cause a pond to fill with sediment, you already know the mechanism by which freshwater succession occurs. Since this process was presented in detail earlier, we will not repeat it here. You should review it as necessary and add it to our list of examples (Figures 4.11 and 4.12)
Climax communities vary in structure
Previously we noted that the organisms present in the climax community on a sand dune are different from those in a forest. It is worth our attention to briefly revisit and expand on that idea here. Keep in mind that time is just one of the many variables affecting ecosystem structure. Moisture, temperature, chemistry, sunlight, and other properties also are very influential. Consequently, even if given thousands of years to develop, a sand dune will never support the organisms that flourish in a temperate forest because its conditions are simply inappropriate for such a community. Similarly, neither tundra nor desert environments will be able to support oak and maple trees. The same is true throughout Earth’s ecosystems: dominant species vary enormously among biomes. Review Table 5.2, above, in this new context to see more examples (the heading on the column labelled “DEFINING ORGANISMS” could also be loosely taken to mean “organisms in a climax community”).
Reality often defies predictions and expectations
The processes described here are idealized representations of what is likely to occur under natural conditions. In other words, early-stage organisms are replaced by later stage-organisms and so forth, but, for numerous reasons, the transitions and progression may not be as predictable or smooth as suggested.
Some organisms resist replacement. Organisms devote a tremendous amount of energy to reproduction and their own continuation. It stands to reason, therefore, that it is not in their best interest to facilitate competitors. Succession occurs because organisms possess different adaptations and abilities to survive under different conditions, it does not result from pioneers willingly or even knowingly serving their fellow members of the biosphere by improving conditions for them. Indeed, some species have developed ways to resist and interfere with later-stage species. A detailed description of the many possible mechanisms by which this can occur is unnecessary for our purposes, but generally speaking, some plants just make it difficult for others to establish themselves. One interesting example of this phenomenon is seen in the way certain grasses form a very thick mat, one that is quite challenging for a seed of a would-be replacer to penetrate. In short, the grasses shield all that newly formed and inviting soil from larger organisms that need it to grow.
Environmental conditions are not static. An ecosystem that is destroyed by a volcano or other event may not be exactly replicated again in the future, even if allowed sufficient time to do so. Environmental conditions like water availability and prevailing temperature can change, favoring different organisms. In addition, as suggested earlier, organisms not native to an area can outcompete those that once dominated. Such invasive species are so well adapted that they can quickly take over, profoundly changing ecosystem structure (more in Chapter 6). Other forces such as disease and nutrient limitations can also influence succession.
Humans affect succession in many ways. Many anthropogenic activities can alter, slow, or even stop ecological succession. A small sampling is provided here, and we will encounter these and other concerns in several places in this textbook, particularly in the chapters found in Part Three.
Ecosystems manipulated for homes, recreation, and sports
Back yards, public parks, golf courses, baseball fields, and similar areas tend to be carefully managed so they are appropriate for their designated uses. Often, they are dominated by grasses that are never replaced by later-stage species. How can this be? The answer is probably familiar to most people: extensive use of lawn mowing, fertilizer, watering, and pesticides help us control the structure of ecosystems (Figure 5.31).

Since it is generally not in the interest of, say, a baseball team, to allow shrubs and ultimately tall trees (assuming the biome in question supports such organisms) to grow in their outfield, conditions are maintained so a grassy field persists indefinitely. In fact, each time you mow your lawn you are preventing succession from occurring—larger plants are simply not given a chance to establish themselves.
Agriculture
If you have ever noticed fields in which crops like corn or wheat are grown, you have seen one way humans stop succession from proceeding (Figure 5.32).

A climax forest might even be cleared from a plot of land, leaving only barren soil behind. Plowing and pesticides help inhibit anything other than the desired crop from growing. We will see much more about this important topic in Chapter 9.
Release of pollutants
Certain biological, chemical, and physical substances are products of industrial, agricultural, domestic (i.e., occur in homes), and military activities. Through numerous mechanisms, these toxins can bring about adverse effects on individuals, populations, communities, and ecosystems. In some cases, succession is stalled because early-stage species such as grasses are better adapted to survive the stress brought about by pollutants than are later-stage ones. One related phenomenon is known as an ecological gradient. In short, dominant species change with distance from a pollution source, providing an important indication that toxic substances are present in an area. Consult Chapter 15 for more on the ways pollutants can affect human and natural systems.
THE CHAPTER ESSENCE IN BRIEF [7]
Ecosystems are made up of the physical and biological components that interact with each other. A variety of interactions are possible and are necessary for organisms to obtain the materials and energy needed to survive and reproduce. Furthermore, ecosystems are not static, but they change with time in response to various stressors.
Think about it some more…[8]
How do organisms use energy? What happens to the energy after they have made use of it?
What would likely happen to heterotrophs (like humans) if autotrophs (like plants and algae) disappeared from Earth?
Is predation always bad for a prey population?
How does the use of antibiotics provide a selective advantage (we also might call it selective pressure) for pathogenic bacteria?
Why would an organism provide a benefit to another? Is that first organism simply generous?
Is it appropriate to refer to your body as an ecosystem?
Why do pioneer species improve conditions for later-stage organisms? Awfully nice of those pioneers, don’t you think?
Given sufficient time, can ecological succession ever convert a desert into a forest ecosystem dominated by oak and maple trees? Why or why not?
- Todar, K. 2016. Online Textbook of Bacteriology, textbookofbacteriology.net. ↵
- Shah, R. 2015. Elements that keep us alive also give color to fireworks. National Institutes of Health. biobeat.nigms.nih.gov ↵
- Karla Moeller. 2013. Boundless biomes. Arizona State University School of Life Sciences Ask A Biologist. https://askabiologist.asu.edu/explore/biomes ↵
- Aq, aquatic or water based; Ter, terrestrial or land based ↵
- Pun intended?? ↵
- For the record: this is a bad idea. ↵
- As you will find throughout this book, here is very succinct summary of the major themes and conclusions of Chapter 5 distilled down to a few sentences and fit to be printed on a t-shirt or posted to social media. ↵
- These questions should not be viewed as an exhaustive review of this chapter; rather, they are intended to provide a starting point for you to contemplate and synthesize some important ideas you have learned so far. ↵
Formally, energy is the capacity to perform work. It enables changes to be made to matter. For example, energy allows you to move a box from the floor to a table. See Chapter 4 for more.
Physical matter that takes up space and has mass. This term is very general and includes many things, for example, individual atoms, chemical compounds, water, soil, and fossil fuels.
One of Earth's four spheres, refers to all living things. See Chapter 4 for more.
One of the known elements in the Universe, carbon (C) atoms consist of 6 protons and 6 electrons. Carbon is an important element for living things; organic C compounds make up the skeleton of biological systems. See Chapter 4 for details.
A chemical element made up of atoms that have 1 proton and 1 electron. It is an important building block for organisms and is the most abundant element in the universe.
A chemical element made up of atoms with 8 protons and 8 electrons. It is an important building block for organisms. Two O atoms bonded together makes up dioxygen, a gas which is essential to the survival of aerobic organisms.
Refers to the many different kinds of microscopic organisms that are single celled and prokaryotic (i.e., do not possess distinct cellular organelles). Collectively, bacteria are extremely diverse, are the oldest organisms on Earth, and are critical to many environmental cycles and processes. See Chapter 3 for more.
The process of converting C from non-biologically available forms to forms that can be used by organisms. Typically, carbon dioxide molecules are converted to glucose. C fixation is carried out by organisms that possess the proper machinery to do so: plants, algae, and some bacteria. Fixation is a general term that indicates an increase in the biological availability of any atom. See Chapter 4 for more details.
Describes the pathways and reservoirs of Earth's water. See Chapters 4 and 11 for more.
Plant-like plankton that are producers; they are consumed by zooplankton. They live in surface waters where sunlight is available. They tend to be on the order of a few millimeters or so in size. See Chapter 5 for more.
A process carried out by certain organisms (plants, algae, and some bacteria) in which the energy of the sun is used to drive the fixation of carbon. See Chapters 4 and 5 for more details.
A group of chemical compounds that include sugars like glucose as well as polymers like starch and cellulose. See Chapter 4 for more.
An organism that eats the flesh of consumers. That is, carnivores usually do not eat primary producers. See Chapter 5 for more.
Refers to organisms that are too small to be seen without the aid of some kind of magnifying lens (e.g., a microscope). Includes bacteria, protozoa, certain forms of algae and fungi, and viruses. See Chapters 1 and 5 for more.
A group of organisms that are microscopic, eukaryotic, mobile (also known as "motile"), and usually heterotrophic. They are roughly ten times larger than bacteria; they also tend to prey on bacteria. See Chapter 3 for more.
Classed as eukaryotic microorganisms. Some are unicellular (e.g., yeast), many are multicellular (e.g., mushroom). They are are heterotrophic and tend to act as important decomposers in ecosystems. See Chapter 3 for more.
Plankton that are consumers or chemoheterotrophs; they eat phytoplankton so live in surface waters. They tend to be several millimeters to a centimeter in size. See Chapter 5 for more.
Remains and waste products of organisms are grouped together as dead organic matter. It contains a lot of stored nutrients that can be used and recycled by decomposers. See Chapters 4 and 5 for more.
An adjective referring to organisms that do not use dioxygen as part of their metabolism. Many such organisms die in the presence of oxygen gas. "Anaerobe" is the noun. Compare to aerobic. See Chapter 1 for more.
An adjective referring to an organism that uses dioxygen in its metabolism. "Aerobe" is the noun, the organism itself. Contrast with anaerobic. See Chapter 1 for more.
A fundamental concept in the science of toxicology. Different species, and even different individuals from the same species, will respond differently to the same poisonous substance. See Chapter 15 for more details.
A general term that includes any disease-causing organism. Some bacteria, fungi, protozoa, and algae are on this list, as are many viruses. Certain worms and other larger organisms can act as pathogens as well. See Chapters 3 and 5 for more.
A larva (plural: larvae) is an immature form of an animal, one that can undergo a dramatic metamorphosis and develop into an adult. For example, caterpillars are butterfly larvae.
A broad term that refers to the number of different organisms in an ecosystem or, often, the whole Earth. There are several specific ways to measure and express biodiversity. See Chapters 5 and 6 for details.
In ecology, organisms that can live on barren landscapes and begin the process of ecological succession. They are pioneer species. See Chapter 5 for more.
